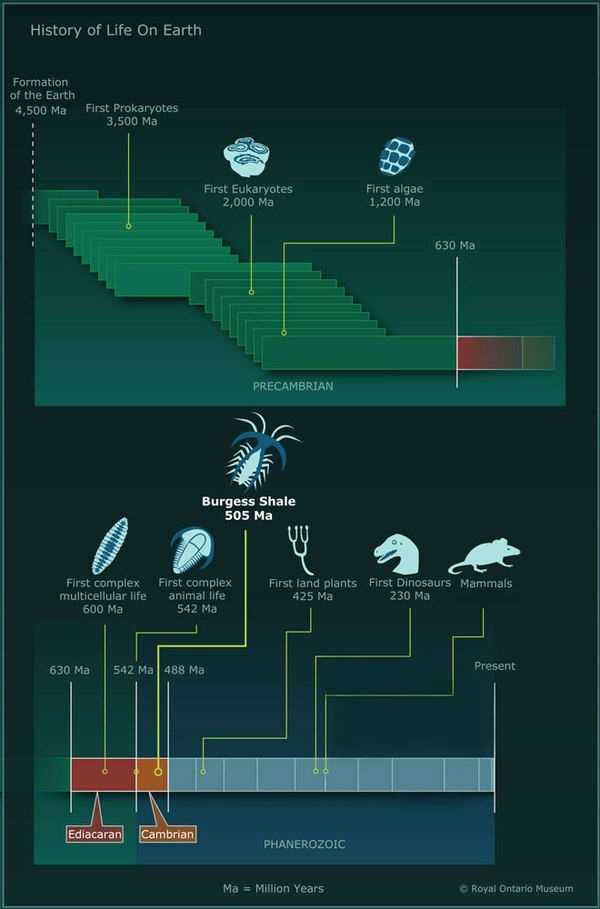
Всем снова привет! В этом посте я постараюсь рассказать об уникальном событии в истории жизни на Земле. Речь пойдет о кембрийском взрыве, или как его еще называют, кембрийской радиации. Кембрийский взрыв - это ярчайшее событие в палеонтологической летописи планеты, когда за относительно небольшой срок (несколько десятков миллионов лет) происходит резкое увеличение количества ископаемых останков живых существ и, на первый взгляд как бы из ниоткуда, появляются почти все современные типы животных (хордовые, членистоногие, моллюски, иглокожие и тд).
У него были предшественники, но Облако удалось захватить идею взрыва кембрия с величайшим красноречием и геологическим утонченность. Помимо его великого руководства и наставничества поколения палеонтологов, он разработал интегративный подход к палеонтологии, добавив навыки в палеогеографию, карбонатную стратиграфию и карбонатную седиментологию. Его более поздняя карьера в Калифорнийском университете, Санта-Барбаре, расширила его интересы до астробиологии и происхождения жизни. Его наблюдения как палеонтолога привели его к тому, чтобы охарактеризовать фанерозойскую летопись окаменелостей как ряд эволюционных извержений, причем кембрий является самым большим из всех.
По времени кембрийский взрыв совпадает с началом кембрийского периода, около 541 млн лет назад. Долгое время феномен резкого увеличения количества и разнообразия ископаемых останков на границе кембрия не имел четкого объяснения. Однако, важность этого события нашла свое отражение во временной шкале истории Земли, разделив геохронологическую шкалу на два больших отрезка: фанерозой - время явной жизни и криптозой или докембрий - время скрытой жизни.
В замечательной, длинной статье, которая привела к этому, он объединил тогда-инновационные исследования, связанные с реконструкцией древнего глобального климата с кембрийским появлением животной жизни. И неужели он действительно эндемичен породам, стратиграфическое положение которых таково, что они не могут быть разумно включены в палеозой? Учитывая данные, он заключил: «Появление многоклеточной животной жизни в кембрии может быть практически столь же неожиданным, как показывает летопись, примером эруптивной эволюции корневого запаса самой эволюции животных».
Ключ к понимаю кембрийского взрыва лежит в структуре самих ископаемых останков. Во всем их разнообразии скрыта одна общая деталь - наличие той или иной формы скелета, будь то раковина моллюска или панцирь членистоногого. А раз у организма есть минерализованные ткани, значит он, скорее всего, будет хорошо сохранятся в ископаемом состоянии, а значит вот мы их и находим. Очевидно, что многие из этих животных жили и раньше, но скелета не имели и в палеонтологической летописи не сохранялись. Следует отметить, что почему-то именно в кембрии часто возникали так называемые лагерштетты - особый тип захоронения окаменелостей, в котором сохраняются даже мягкие ткани животных. Самые знаменитые лагерштетты - это Сланцы Бёрджес в Канаде.
Клауд подчеркнул необходимость найти связь между изменением глобальной окружающей среды и эволюцией извержения кембрия. Его первоначальный акцент на растворенном кислороде, основанный на доказательствах из полосчатых железных образований, не выдержал испытания временем; но его акцент на доказательствах был краеугольным камнем исследований Кембрия.
Более поздние крупные палеонтологические открытия тесно связаны с изучением филогенетических отношений ранних билатеровых групп с большими уточнениями геологической шкалы. Самые последние геологические временные масштабы показывают значительное улучшение, и текущая оценка начала кембрия на уровне 542 млн. Лет, скорее всего, с точностью до миллиона лет.
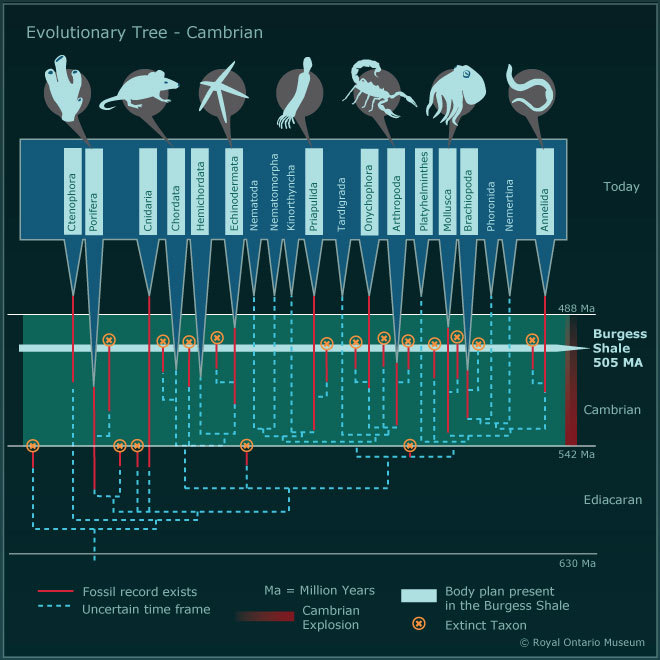
Причина такого единовременного обретения скелетов разными типами животных до конца не ясна. Есть несколько гипотез, большинство из которых связывают обретение скелета с ростом содержания кислорода в мелководных морях, где и обитал почти весь кембрийский зоопарк. На суше в то время не было почвы, а поверхность континентов представляла собой большей частью либо голые скалы, либо песчаные россыпи.
Свидетельства о происхождении и расхождении двухсторонних
Свидетельства о происхождении и расхождении билатерианцев подразделяются на четыре категории: данные молекулярных часов, данные об ископаемых по появлению билатеровых групп, морфологическое и филогенетическое исследование летописи окаменелостей; и генетические доказательства.
Объяснения на основе развития организмов
Молекулярные часы основываются на презумпции отношения между временем, прошедшим с того момента, когда две линии расходятся, и степенью генетической разницы между ними, основанной на идее интеграции эволюционной скорости со временем. Время можно оценить, взяв две сестринские эволюционные линии, А и В, и найти датированные окаменелости каждой группы. Если летопись окаменелостей была полна, а генетическая дивергенция была одинаковой постоянной для всех генов за время, прошедшую после раскола, этот процесс был бы легким.
![]()
Одна из правдоподобных гипотез связана с появлением эффективных морских фильтраторов. Они, очищая воду от органической взвеси, делали ее более прозрачной для солнечного света, благодаря чему росло содержание фитопланктона, который в свою очередь производил больше кислорода и сам по себе являлся пищей. Рост кислорода сделал энергетически выгодным создание скелета и мышц, что позволило появиться многоклеточным хищникам и запустило эволюционную гонку "хищник-жертва".
Можно было бы установить расхождения в расхождениях, и тогда можно было бы экстраполировать этот показатель, чтобы объяснить время расхождения для двух родословных, которые являются древними и чьи даты ископаемого происхождения неизвестны. Эти предубеждения привели к ряду исследований, которые пытаются исправить неоднородность скорости в разных частях дерева или просто отказаться от тех случаев, когда такая гетерогенность существует. В некоторых исследованиях было предпринято попытку включить большое количество генов, которые могли бы усреднить вариацию по закону центральной тенденции.

Начинается все кембрийское разнообразие с так называемой фауны мелких ракушек, массово встречающейся в начале кембрия. Эта фауна представляет собой разнообразные небольшие скелетные элементы: чешуйки, спикулы, трубочки, ракушки. В большинстве случаев непонятно, какому животному они могли бы принадлежать. Ведь многие группы животных, которые в наше время имеют монолитный скелет (например, моллюски) изначально были мягкотелыми, потом покрывались отдельными скелетными элементами (спикулами, ракушками), и только потом эти элементы срастались в единое целое (раковину).
«Гонка вооружений» между хищниками и жертвами
Все основные исследования последовательно дают дату расхождения для протостомов и дейтеростомов значительно до начала кембрия. Более поздние исследования использовали больше генов, но дали широкий диапазон дат. Однако последовательно эти даты являются докембрийскими. Если бы все основные филы расходились за очень короткий промежуток времени, мы могли бы ожидать проблем в реконструкции, которые возникли, поскольку близко расположенные узлы в сотни миллионов лет были бы почти невозможны для решения.
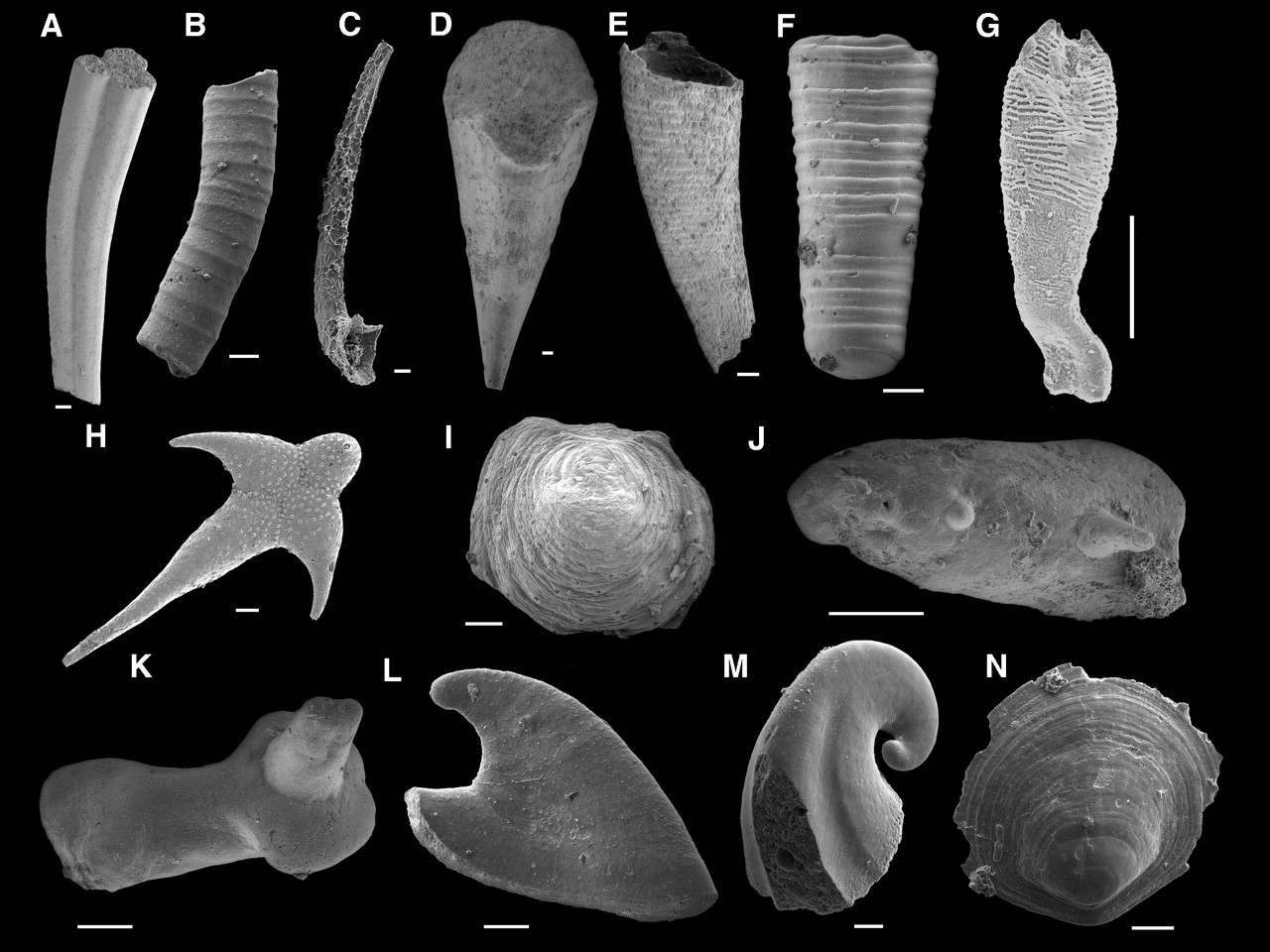
Кроме фауны мелких ракушек, кембрийский взрыв являет на свет уже "готовых" моллюсков, брахиопод, губок, трилобитов и пр., переходные формы между разными типами животных, а также массу удивительных, словно из фантастического фильма, существ. У животных возникает современный план строения нервной системы, развиваются органы чувств, прежде всего глаза.
По-прежнему существуют большие разногласия в отношении методов и подходов. Более ранние исследования с использованием нескольких генов были подвергнуты критике за включение нескольких генов со слишком высокой степенью гетерогенности ставок по деревьям, но в результате замещающих исследований критиков не удалось установить даты, согласующиеся с кембриями, и также не смогли создать даты, которые очень совместимы между генами. Петерсон и его коллеги пришли к выводу, что в предыдущих исследованиях использовались гены с темпами эволюции, которые не подходят для изучения большей части филы, участвующих в кембрийском взрыве, но их коррекция все же дала даты, предшествующие кембрии, на 30 миллионов до 114 миллионов лет.

Причудливые животные кембрия: семиглазая опабиния с хоботком, заканчивающимся клещней; галлюциногения с множеством гибких ножек и шипами на спине; хордовое пикайя с парой антенн-щупалец на голове; маррелла с мощным роговым щитом с четырьмя отростками; айшеайа - онихофор с телескопическими ножками.
Блэр и Хеджес пересмотрели эту самую ювенильную оценку расхождений между кембрийскими билатерами и обнаружили, что они в основном получены из набора ископаемых калибровок, которые приводят к искажениям в сторону более медленных расхождений, а не к различным темпам молекулярного расхождения между позвоночными и беспозвоночными, как утверждает Петерсон и коллег. С быстрым увеличением эволюции последовательности в начале излучения регулярные постоянные молекулярные часы могут переоценить время расхождения.
Но выбор наивысшей скорости эволюции последовательности по-прежнему будет увеличивать время расхождения до 586 млн. Лет назад. Таким образом, хотя большой диапазон оценок времени дивергенции не внушает доверия, мы все равно должны столкнуться с текущим заключением о том, что молекулярные оценки не совпадают с данными о происхождении окаменелостей, что создает большую радиацию между приблизительно 540 млн.
Иногда принадлежность ископаемого к тому или иному животному бывает очень трудно определить, и только нахождение новых, более полных и подробных отпечатков, приоткрывает эту тайну.

Загадка аномалокариса. Найденные по отдельности опечатки его частей тела поначалу трактовались как отдельные животные. Ротовые придатки - как какие-то креветки, ротовое отверстие - как медуза странной формы, а тело - вообще как морская губка.
Лет и 520 млн лет назад. Данные окаменелостей поддерживают вывод, противоречащий оценкам молекулярных часов. Билатеровые группы животных, по-видимому, появляются в летописи окаменелостей или непосредственно перед началом кембрия. За последние 15 лет в породах, относящихся к раннему кембриям, были обнаружены члены все большего числа фила и крупных классов билатеров, включая позвоночных. Буквальное толкование летописи окаменелостей может привести к полному расхождению билатеров примерно за 20 миллионов лет или меньше.
Следы окаменелостей, которые являются норами и следами, зарегистрированными в отложениях, появляются в результате взрыва вблизи основания кембрия. В значительной степени из этого разнообразия преобладают норы, которые служили приютами, из которых инфаунальные животные питались или двигались к поверхности осадка, но следы членистоногих также становятся заметными в раннем кембрии. Возникновение биотурбации в конце позднего протерозоя, возможно, было причиной разрушения микробных матов, которые доминировали над поверхностью осадка в Эдиакаране наряду с некоторыми горизонтальными следами.
Толщу кембрийских морей бороздили и другие гиганты - тамисиокарисы. Они были родственны аномалокарису, однако питались по-другому, фильтруя воду своими приротовыми конечностями, загибая их, и отправляли отфильтрованный комок пищи к круглому рту. Основу их рациона составлял разнообразный зоопланктон. Тамисиокарисы были неким аналогом современных китовых акул, которые тоже отказались от хищничества. Наличие такого фильтратора как тамисиокарис говорит о том, что уже раннем кембрии в морях была полно представлена пищева цепочка: от фитопланктона и зоопланктона к крупным фильтраторам и хищникам.
Глубокий рыть, вероятно, ответ на поверхностных хищников, плохо зарегистрирован до ордовика и даже позже. Всегда остается открытым вопрос о том, является ли кажущееся внезапное появление двухсторонних людей в раннем кембрии результатом сохранения бреши. Вполне возможно, что камни, не соответствующие сохранению, доминируют во времени до кембрия. Эдиакаранские окаменелости в последней части протерозоя в изобилии, но их обычно встречают в песке, который не сохранит тонкие структуры, наблюдаемые в органических пленках, обнаруженных в более мелкозернистых отложениях нижнего кембрия.

Тайна кембрийского взрыва, когда как бы из ниоткуда в ископаемом состоянии появляются многие современные типы животных, что должно было бы явственно свидетельствовать об Акте творения, при детальном взгляде оказывается не такой уж тайной. Кембрийский взрыв, а вернее Кембрийская скелетная революция, стала итогом многомиллиардолетней эволюции жизни на Земле, когда одновременно появилась возможность и возникла необходимость перейти на более высокий уровень метаболизма, а парадигма "хищник-жертва" во много раз ускорила темп эволюции и открыла новые возможности строения животных.
С отсутствием растворенного кислорода - или, возможно, минерализованных скелетов - до кембрия билатерианцы могли быть довольно маленькими по размеру тела, что уменьшало бы вероятность сохранения. Конечно, в докембрийских породах моложе 750 Ма до 850 Мах особый особый тип деликатной консервации Берджес-Сланца отсутствует, что оставляет значительный промежуток времени до появления раннего кембрия.
Морфология и филогенетика, обнаруженные окаменелостями
Возможно, самым сильным свидетельством поддержки эволюционного взрыва кембрийских форм животных является первое явное появление в раннем кембрии скелетных окаменелостей, представляющих представителей многих морских животных билатерианских животных. Впечатление от взрыва усиливается рядом окаменелостей с неясной близостью к сохранившейся филе. Сначала утверждалось, что ранний кембрий изобилует формами, которые не имеют очевидного сходства с сохранившейся филой или даже с другими древними группами.
Кембрийский взрыв - внезапное (в геологическом смысле) появление в раннекембрийских (около 540 миллионов лет назад) отложениях окаменелостей представителей многих подразделений животного царства, на фоне отсутствия их окаменелостей или окаменелостей их предков в докембрийских отложениях. По мере накопления палеонтологических данных интерпретация кембрийского взрыва неоднократно изменялась.
Некоторые виды имеют символы, которые могут помещать их в качестве предковых членов сохранившейся филы, но существуют разногласия в отношении таких групп, как халкиреиды. Известный таксономический уклон подкрался в исследования кембрийских и других ранних животных окаменелостей. Когда была обнаружена странная окаменелость, неклассифицируемые части тела влияли на палеонтологов, чтобы классифицировать такие организмы как членов новых классов сохранившихся фил или даже новых фил. По иронии судьбы, это как раз противоположность тому, что утверждал Гулд, - это неудача великого палеонтолога Уолкотта, который якобы имел тенденцию объединять самые странные организмы в обычных группах, которые уже были описаны.
История и проблематика
В целом ранняя проблематика «кембрийского взрыва» состояла в попытках объяснить внезапное появление окаменелостей на нижней границе кембрия и их отсутствие в более древних отложениях - как свидетельствах существования или несуществования жизни в предшествующий период истории Земли .
Возможно, Гулд был прав относительно Уолкотта, но он пропустил остальную часть картины. С гей-отбросом палеонтологи называли ранние таксоны животных и определяли их как членов новой филы или классов. По сути, палеонтологи вознаграждаются признанием за открытие нового таксона, когда они назначают его на более высокий уровень классификации. Тенденция ускорилась вторым исследованием Бургесского сланца Гарри Уиттингтоном и его коллегами. Странная, колючая, червеобразная окаменелость была причудливо названа Галлюцигения и считалась таксоном, не связанным с обычной известной филой.
Современный интерес к данной теме был подогрет работой Гарри Уиттингтона (англ. ) и его коллег, которые в 1970-х годах повторно проанализировали ряд окаменелостей из сланцев Бёрджес и заключили, что большинство из них являются останками организмов, принципиально отличных от любых ныне существующих животных (см. вендобионты) . Ставшая популярной работа Стивена Гулда «Удивительная жизнь» открыла эту тему для широкой публики и вновь сделала актуальной проблему существования и природы кембрийского взрыва. Хотя и существенно расходясь в деталях, как Уиттингтон, так и Гулд предположили, что все современные типы животного царства возникли на границе кембрия почти внезапно, не будучи потомками ранее существовавшей фауны.
Еще одна ископаемая, ранее считавшаяся Уолкоттом как аннелид, была повторно описана как принадлежащая к новому типу, возможно, связанному с моллюсками. Этот уклон заставил понятие эволюционной лужайки, в которой многочисленные бессознательные таксоны внезапно появились в кембрии, которые прекрасно сочетались с концепцией Облака о полифилетическом происхождении животной филы. Два важных прорыва изменили научную концепцию кембрийского взрыва как эволюционный газон странных и несвязанных побегов: пересмотр морфологии этих «странных»существ и переосмысление этих разрозненных таксонов как членов эволюционного дерева, которое представляет морфологические признаки различные группы с точки зрения эволюционной связанности.
Однако другие исследования, как относящиеся к 1970-м годам, так и более поздние, отмечали, что сложные животные, сходные с современными видами, возникли задолго до начала кембрия. В любом случае, многочисленные свидетельства в пользу существования жизни задолго до кембрия сняли с повестки дня вопрос о «кембрийском взрыве», как феномене внезапного возникновения жизни.
Вопрос же о причинах и механизмах очередного роста сложности и разнообразия форм жизни на границе кембрия, не являющихся потомками исчезнувших хайнаньской и вендской биот , остаётся открытым. На сегодняшний день проблематика «кембрийского взрыва» сосредоточена на двух ключевых вопросах:
- действительно ли в раннем кембрии произошёл «взрывной» рост разнообразия и сложности организмов, и
- что могло послужить причиной столь быстрой эволюции.
Источники информации
Построение точной хронологии событий на границе между докембрием и ранним кембрием представляет значительную трудность. В силу этого описания последовательности и взаимосвязи тех или иных явлений в рамках обсуждения кембрийского взрыва следует рассматривать с определённой осторожностью.
Помимо проблем с датировкой, изучение событий на границе кембрия затруднено нехваткой самого палеонтологического материала. К сожалению, чем дальше отстоит от нас исследуемый период, тем менее доступны для изучения его окаменелости. Среди прочего, причинами этого являются:
Останки живых организмов
Наряду с окаменелостями, кембрийские отложения содержат необычно высокое число месторождений, сохранивших отпечатки мягких частей тел различных организмов. Такие отпечатки позволяют детально изучить животных, которые не сохраняются в виде окаменелостей, а также внутреннее устройство и функционирование организмов, которые обычно представлены только раковинами, шипами, когтями и т. д.
В кембрии наиболее важными месторождениями являются: раннекембрийские
среднекембрийские
и верхнекембрийское месторождение
- Орстен (Швеция).
Хотя все эти месторождения превосходно сохранили анатомические детали организмов, и они далеки от совершенства. Большая часть кембрийской фауны, возможно, вообще в них не представлена, поскольку месторождения формировались в специфических условиях (оползнях или вулканическом пепле, очень быстро сохранявших мягкие части тел). Вдобавок к этому, известные месторождения покрывают лишь ограниченный период кембрия и не затрагивают важнейшее время непосредственно перед его началом. Поскольку хорошо сохранившиеся захоронения вообще редки, а месторождения окаменелостей - крайне редки, весьма маловероятно, что в них представлены все типы существовавших организмов.
Окаменелые следы, оставленные живыми организмами
Окаменелые следы состоят, главным образом, из дорожек и норок, оставленных на морском дне. Такие следы крайне важны, поскольку они предоставляют исследователю данные об организмах, чьи тела не сохранились в окаменелостях. Зачастую только они позволяют изучить организмы, относящиеся к периоду, от которого не сохранились останки животных, способных оставить такие следы. Хотя точное соотнесение следов с оставившими их организмами, как правило, невозможно, следы могут служить наиболее ранним доказательством существования относительно сложных животных (подобных, например, дождевым червям).
Геохимические наблюдения
В геологических породах, относящихся к нижней границе кембрия и его началу, отмечены сильные колебания изотопного состава трех элементов - стронция (87 Sr/ 86 Sr), серы (34 S/ 32 S) и углерода (13 C/ 12 C).
- Массовое вымирание . Массовые вымирания организмов должны непосредственно повышать долю изотопа 12 C в составе отложений и таким образом уменьшать соотношение 13 C/ 12 C. Следующее за вымиранием падение продуктивности «биологического насоса» вновь снижает концентрацию 12 C в геологических породах.
- Высвобождение метана . В вечной мерзлоте и на континентальном шельфе вырабатываемые бактериями молекулы метана удерживаются в «клетке» из молекул воды, формируя смесь, называемую клатрат метана . Будучи произведен живыми организмами, этот метан обогащен изотопом 12 C. По мере роста температуры или падения атмосферного давления, клатраты распадаются. Подобный распад высвобождает в атмосферу запасенный метан, обогащенный углеродом-12. В атмосфере метан преобразуется в диоксид углерода и воду, причем диоксид углерода взаимодействует с минералами, формируя углеродсодержащие породы с избытком углерода-12. В результате изотопный состав геологических отложений смещается в сторону 12 C.
Сравнительная анатомия
Кладистика - это метод построения «эволюционного дерева» организмов, чаще всего - путем сопоставления их анатомического строения. При помощи такого анализа можно сопоставлять между собой как современные, так и ископаемые организмы для установления хода их эволюции. В ряде случаев можно заключить, что группа A должна была появиться раньше групп B и C, поскольку те имеют больше сходства между собой, чем с A. Сам по себе (без соотнесения с данными палеонтологических раскопок) этот метод ничего не говорит о времени, когда происходили изменения, однако он способен восстанавливать последовательности эволюционного развития организмов.
Молекулярная филогенетика
Палеонтологические свидетельства
В этом разделе основные свидетельства упорядочены по времени формирования тех отложений, в которых они были обнаружены, поскольку датировка является центральным вопросом при изучении кембрийского взрыва. При этом следует помнить о неоднозначности хронологии ископаемых, относящихся к этому периоду.
Обзор находок начинается со времени, задолго предшествующего кембрию, и заканчивается в раннем ордовике, поскольку существует мнение, что формирование основных типов современной фауны началось до и закончилось после кембрия.
Данные молекулярной филогенетики (1,2 - 0,5 млрд лет)
До сих пор продолжаются дебаты относительно хронологической интерпретации данных молекулярной филогенетики:
В любом случае, данные молекулярной филогенетики говорят о том, что формирование основных типов животных было весьма длительным процессом, далеко выходящим за рамки 10 млн лет (около 543-533 млн лет назад) «кембрийского взрыва».
Докембрийские следы многоклеточных
Существуют как косвенные, так и прямые палеонтологические свидетельства того, что многоклеточные организмы впервые возникли задолго до начала кембрия.
Упадок бактериальных матов (1,25 млрд лет)
Докембрийский строматолит
![]()
Современные строматолиты. Западная Австралия.
Строматолиты составляют важную часть окаменелостей, начиная со времени около 3 млрд лет назад. Их расцвет приходится на период в 1,25 млрд.лет назад, после чего они стали приходить в упадок (как в общей численности, так и в разнообразии). К началу кембрия такое сокращение составило уже около 20 %.
Наиболее распространенным объяснением упадка является предположение о том, что микроорганизмы, входящие в состав бактериальных матов, стали жертвами поедания («обгрызания строматолитов») со стороны других живых организмов (что должно свидетельствовать о существовании довольно сложных хищников уже около 1 млрд лет назад). Это предположение подтверждается наблюдаемыми антикорреляциями между разнообразием и численностью строматолитов - с одной стороны и богатством морской фауны - с другой. Так, повторный упадок строматолитов пришёлся на поздний ордовик - сразу после очередной «вспышки» разнообразия и численности морской фауны. В ходе ордовикско-силурийского и пермско-триасового вымираний вновь наблюдалось восстановление строматолитов - с последующими упадками по мере восстановления морской фауны.
Развитие средств защиты у акритархов. Раннее хищничество (1 млрд лет)
Акритархи - это окаменелые остатки ископаемых неопределенной природы, как правило - оболочки цист одноклеточных и многоклеточных водорослей . Впервые они обнаруживаются в отложениях, датируемых временем 2 млрд лет назад.
Около 1 млрд лет назад произошёл резкий рост в их численности, разнообразии, размерах, анатомической сложности и, особенно, в количестве и видах колючек. Численность архитархов резко сократилась в ходе глобального оледенения , однако впоследствии восстановилась с достижением максимального разнообразия уже в палеозое .
Их исключительно колючие формы, относящиеся к периоду 1 млрд лет назад, могут свидетельствовать о существовании уже тогда хищников, достаточно крупных, чтобы раздавливать их или заглатывать целиком. Другие группы небольших организмов неопротерозоя также имеют те или иные средства защиты от хищников.
Следы, оставленные многоклеточными организмами (1 млрд лет)
В Индии в составе отложений, датируемых 1 млрд лет назад, обнаружены окаменелости, которые могут быть следами организмов, двигавшихся через и сквозь мягкие породы. Найденные следы оставлены, очевидно, непосредственно под слоем цианобактериальных матов , покрывавших морское дно. Исследователи заключили, что следы обязаны своим появлением перистальтике трехслойных многоклеточных размером до 5 мм - иными словами, животными, чей диаметр был сравним с диаметром земляных червей, и возможно, имевшими целом . Другие исследователи считают, что эти и подобные им находки старше 600 млн лет, были оставлены не живыми организмами, а физическими процессами.
Многоклеточные эмбрионы из Доушаньто (632-550 млн лет)
Однако обнаружение в 2007 году эмбрионов, окружённых сложно устроенной оболочкой (в породах возрастом 580-550 млн лет), свидетельствует о том, что окаменелости в Доушаньто - не что иное, как покоящиеся яйца многоклеточных беспозвоночных . Более того, стало ясно, что часть акритархов, обнаруженных в более ранних породах Доушаньто (632 млн лет), в действительности представляют оболочки таких эмбрионов
Другое ископаемое из Доушаньто - Vernanimalcula (от 0,1 до 0,2 мм в диаметре, возраст около 580 млн лет) - рассматривается рядом ученых как останки трехслойного билатерального организма, имевшего целом , то есть животного столь же сложного, как дождевые черви или моллюски . Несмотря на сомнения в органической природе этих ископаемых , поскольку все 10 найденных образцов Vernanimalcula имеют одинаковые размер и конфигурацию, маловероятно, чтобы подобное однообразие было результатом неорганических процессов.
В наиболее поздних отложениях Доушаньто также обнаружено резкое падение соотношения изотопов углерода 13 C/ 12 C. Хотя это изменение является общемировым, оно не совпадает по времени с другими крупными событиями, такими как массовые вымирания . Возможное объяснение состоит в «цепной реакции» взаимосвязанной эволюции организмов и изменений химического состава морской воды. Многоклеточные организмы, активно поглощая углерод из воды, могли способствовать росту концентрации кислорода, растворенного в морской воде, в свою очередь, обеспечивая появление новых многоклеточных (таких как Namapoikia ).
Эдиакарская фауна (610-543 млн лет)



Сприггина
Многоклеточные ископаемые животные эдиакарского периода были впервые обнаружены в местонахождении Эдиакарские холмы в Австралии, а затем и в отложениях из других регинов: Чарнвудский лес (Англия) и полуостров Авалон (Канада). Эти ископаемые имеют возраст 610-543 млн лет (эдиакарский период предшествует кембрийскому). Большая их часть имела размеры в несколько сантиметров и была существенно крупнее своих предшественников. Многие из этих организмов не имеют аналогов ни с одним из видов, обитавших до или после эдиакарского периода. Высказывались предположения, что наиболее «странные» представители эдиакарской фауны должны быть отнесены к отдельному царству - «Вендобионты» (Vendozoa). Именно к их числу относится чарния - наиболее древняя из находок эдиакарского периода (возраст - 580 млн лет).
Однако некоторые эдиакарские организмы могут оказаться предшественниками более поздней фауны:
Отверстия в раковинах Cloudina . Отбор в системе «хищник-жертва»
В ряде мест до 20 % окаменелостей Cloudina содержат отверстия диаметром от 15 до 400 мкм, оставленные хищниками. Некоторые Cloudina были повреждены несколько раз, что говорит об их способности переживать атаки (хищники повторно не атакуют пустые раковины). Очень схожие с Cloudina ископаемые Sinotubulites , обнаруживаемые в тех же захоронениях, вообще не содержат отверстий. Такая избирательность может свидетельствовать о существовании уже в эдиакарском периоде эволюционного отбора размерных классов, а также специализации жертв в ответ на хищничество, рассматриваемой как одна из причин кембрийского взрыва.
Рост разнообразия следов, оставленных организмами (565-543 млн лет)
Наиболее ранние эдиакарские ископаемые, относящиеся ко времени 610-600 млн.лет назад, содержали только следы, оставленные стрекающими . Около 565 млн лет назад появляются более сложные следы. Чтобы их оставить, организмам требовался кожномускульный мешок , а их общее строение - быть сложнее, чем у стрекающих или плоских червей .
Непосредственно перед началом кембрия (около 543 млн лет назад) возникает множество новых следов, включая вертикальные норки Diplocraterion и Skolithos ), а также следы возможных членистоногих (Cruziana и Rusophycus ). Вертикальные норки свидетельствуют о том, что червеподобные животные приобрели новое поведение и, возможно, новые физические возможности. Следы же Cruziana и Rusophycus говорят о существовании у непосредственных предшественников членистоногих экзоскелета , хотя возможно и не столь жесткого, как впоследствии.
Кембрийские ископаемые
Раковинная фауна (543-533 млн лет)
Ископаемые, известные как «мелкая раковистая фауна» (англ. small shelly fossils ), были обнаружены в разных частях света и датируются концом венда (немакит-далдынский ярус) и первыми 10 млн лет с начала кембрия (томмотский ярус). К ним относится весьма разнообразная коллекция ископаемых: иглы, склериты (пластинки брони), трубки, археоциаты (группа губок или близких к ним животных), а также небольшие раковины, весьма напоминающие брахиопод и улиткоподобных моллюсков , хотя и очень маленьких (1-2 мм в длину).
Раннекембрийские трилобиты и иглокожие (530 млн лет)
Фауна Сириус Пассет (527 млн лет)
Наиболее распространенными ископаемыми гренландского захоронения Сириус Пассет являются членистоногие. Также имеется некоторое число организмов с твердыми (минерализованными) частями тела: трилобиты , хиолиты , губки , брахиоподы . Полностью отсутствуют иглокожие и моллюски .
Наиболее странными организмами Сириус Пассет были Pambdelurion и Kerygmachela . Их длинное тело, состоящее их мягких сегментов, с парой широких «плавников» на большинстве сегментов и парой сегментированных придатков сзади делают их схожими с аномалокаридами . При этом наружные части верхней поверхности «плавников» имели рифлёные поверхности, которые могут быть жабрами . Под каждым «плавником» имеется короткая бескостная нога. Такое строение позволяет связать их с членистоногими .
Чэнцзянская фауна (525-520 млн лет)

Haikouichthys - реконструкция

Аномалокарис - реконструкция

Hallucigenia - реконструкция
Эта фауна описана из нескольких мест обнаружения окаменелостей в уезде Чэнцзян (городской округ Юйси , провинция Юньнань , Китай). Наиболее важным является Maotianshan shale - захоронение, в котором очень хорошо представлены окаменелости мягкотелых животных. Чэнцзянская фауна относится к периоду 525-520 млн лет назад - середине раннего кембрия, на несколько миллионов лет позднее Sirius Passet и, по меньшей мере, на 10 млн лет раньше сланцев Бёрджес.
В окаменелостях обнаружены части тела древнейших хордовых (тип, к которому относятся все позвоночные):
В этих же отложениях найдены представители групп, близких к членистоногим:
Вероятно, эти организмы относятся к группе Lobopodia , к которой из современных групп принадлежат онихофоры .
Около половины чэнцзянских ископаемых являются членистоногими , часть которых обладала твердым, минерализованным экзоскелетом , как и большинство более поздних морских членистоногих. Лишь 3 % организмов обладали твердыми панцирями (в основном - трилобиты). Здесь также обнаружены представители многих других типов животных :
- Приапулиды (роющие морские черви - засадные хищники);
- Щетинкочелюстные (морские беспозвоночные, входящие в состав планктона);
- Гребневики (кишечнополостные, внешне похожие на медуз);
- Иглокожие (морские звезды , морские огурцы и т. п.),
- Хиолиты (загадочные животные, имевшие маленькие конические раковины),
Раннекембрийские ракообразные (520 млн лет)
Сланцы Бёрджес (515 млн лет)
Основная статья: Бёрджес-Шейл
 Marrella
Marrella


Pikaia - реконструкция
Сланцы Бёрджес - первое из известных крупных захоронений кембрийского периода, открытое Уолкоттом в 1909 году. Повторный анализ ископаемых, выполненный Уиттингтоном и его коллегами в 1970-х, лег в основу книги Гулда «Удивительная жизнь», открывшей кембрийский взрыв для широкой публики.
Среди ископаемых сланцев Бёрджес наиболее распространены членистоногие , однако многие из них необычны и с трудом поддаются классификации:

Opabinia - реконструкция

Wiwaxia - реконструкция
Кроме того, в захоронении представлены образцы экзотических организмов:
Возникновение новых экосистем и типов после кембрия
Из-за крупного вымирания на границе кембрия и ордовика типичные для палеозоя морские экосистемы сформировались лишь в ходе последующего восстановления морской фауны. Наиболее ранние ископаемые, относящиеся к мшанкам , также впервые обнаруживаются в ордовикском периоде - много позже «кембрийского взрыва».
Выводы
Длительный процесс возникновения многоклеточных
Во времена Дарвина все, что было известно об ископаемых останках, позволяло предположить, что основные типы многоклеточных возникли и сформировались в течение всего нескольких миллионов лет - от раннего до среднего кембрия. Вплоть до 1980-х годов эти представления ещё оставались в силе.
Однако последние находки свидетельствуют о том, что по меньшей мере некоторые трехслойные билатеральные организмы существовали до начала кембрия: Kimberella можно рассматривать как ранних моллюсков, а царапины на скальных породах возле этих ископаемых позволяет предположить моллюскоподобный же метод питания (555 млн.лет назад). Если же предположить, что Vernanimalcula имела трехслойный билатеральный целом , это отодвигает возникновение сложных животных ещё на 25-50 млн.лет назад. Обнаружение отверстий в оболочке Cloudina также позволяет предположить наличие развитых хищников в конце эдиакарского периода. Кроме того, некоторые следы в ископаемых, относящихся к середине эдиакарского периода (около 565 млн лет назад), могли быть оставлены животными, более сложными, чем плоские черви и имеющими кожномускульный мешок.
Задолго до этого длительный закат строматолитов (начавшийся около 1,25 млрд лет назад), говорит о раннем возникновении животных, достаточно сложных для их «обгрызания» . Рост же изобилия и разнообразия шипов у акритархов в это же время приводит к выводу, что уже тогда существовали хищники, достаточно крупные для того, чтобы такая защита была необходима. На другом конце временной шкалы, относящейся к кембрийскому взрыву, нужно отметить отсутствие ряда основных типов нынешней фауны вплоть до конца кембрия, а типичных палеозойских экосистем - вплоть до ордовика .
Таким образом, на сегодня опровергнута точка зрения, согласно которой животные «современного» уровня сложности (сравнимого с ныне живущими беспозвоночными) возникли в течение всего нескольких миллионов лет раннего кембрия. Тем не менее, подавляющая часть современных типов впервые появилась в кембрии (за исключением моллюсков , иглокожих и членистоногих , возможно, возникших в эдиакарский период). Кроме того, взрывной рост таксономического разнообразия также наблюдался в начале кембрия.
«Взрыв» таксономического разнообразия в раннем кембрии
«Таксономическое разнообразие» означает количество организмов, существенно различающихся по своему строению. В то же время «морфологическое разнообразие» означает общее число видов и ничего не говорит о числе базовых «дизайнов» (возможно множество вариаций небольшого количества основных типов анатомического строения). Нет сомнений, что именно таксономическое разнообразие резко возросло в раннем кембрии и сохранялось на этом уровне в течение всего периода - мы можем обнаружить современно выглядящих животных (таких как ракообразные , иглокожие , и рыбы) почти в одно и то же время, а зачастую - и в общих захоронениях с организмами типа аномалокариса и Halkieria , которые считаются «дядями» или «пра-дядями» современных видов.
При более близком изучении обнаруживается другой сюрприз - некоторые современно выглядящие животные, например раннекембрийские ракообразные , трилобиты и иглокожие , находятся в более ранних отложениях, чем некоторые «дяди» или «пра-дяди» ныне живущих групп, не оставившие прямых потомков. Это может являться результатом разрывов и вариаций формирования ископаемых отложений, либо означать, что предки современных организмов эволюционировали в разное время и, возможно, с разной скоростью.
Возможные причины «взрыва»
Несмотря на то что довольно сложные трехслойные животные существовали до (а возможно, задолго до) кембрия, эволюционное развитие в раннем кембрии представляется исключительно быстрым. Предпринималось множество попыток объяснить причины подобного «взрывного» развития.
Изменения окружающей среды
Рост концентрации кислорода
Самая ранняя атмосфера Земли вообще не содержала свободного кислорода. Тот кислород, которым дышат современные животные - как содержащийся в воздухе, так и растворенный в воде - является продуктом миллиардов лет фотосинтеза , главным образом - микроорганизмов (таких как цианобактерии). Примерно 2,5 миллиарда лет назад концентрация кислорода в атмосфере резко возросла. До этого времени весь вырабатываемый микроорганизмами кислород полностью тратился на окисление элементов с высоким сродством к кислороду, таких как железо. Пока не произошло их полное связывание на суше и в верхних слоях океана, в атмосфере существовали лишь локальные «кислородные оазисы».
Нехватка кислорода могла длительное время препятствовать развитию крупных сложных организмов. Проблема состоит в том, что количество кислорода, которое животное может абсорбировать из окружающей среды, ограничено площадью поверхности (легких и жабр у наиболее сложных животных; кожи - у более простых). Количество же кислорода, требуемое для жизнедеятельности, определяется массой и объёмом организма, которые по мере увеличения размеров растут быстрее, чем площадь. Рост концентрации кислорода в воздухе и в воде мог ослаблять или вовсе устранять это ограничение.
Нужно отметить, что достаточное количество кислорода для существования крупных вендобионтов присутствовало уже в эдиакарский период. Однако дальнейший рост концентрации кислорода (между эдиакарским и кембрийским периодами) мог предоставить организмам дополнительную энергию для производства веществ (таких как коллаген), необходимых для развития принципиально более сложных структур тела, в том числе - используемых для хищничества и защиты от него.
Земля-снежок (Snowball Earth)
Существуют многочисленные доказательства того, что в позднем неопротерозое (включая ранний эдиакарский период) Земля подвергалась глобальному оледенению, в ходе которого большая часть её была покрыта льдом, а температура поверхности была близка к точке замерзания даже на экваторе. Некоторые исследователи указывают, что это обстоятельство может быть тесно связано с кембрийским взрывом, поскольку самые ранние из известных ископаемых относятся к периоду вскоре после конца последнего полного оледенения.
Однако довольно трудно указать причинно-следственную связь таких катастроф с последующим ростом размеров и сложности организмов. Возможно, низкие температуры увеличивали концентрацию кислорода в океане - его растворимость в морской воде растет почти вдвое при падении температуры с 30 °C до 0 °C.
Флуктуации изотопного состава углерода
В отложениях на границе эдиакарского и кембрийского периодов наблюдается очень резкое снижение, а вслед за ним - необычно сильные колебания соотношения изотопов углерода 13 C/ 12 C в течение всего раннего кембрия.
Многие ученые предполагали, что исходное падение связано с массовым вымиранием непосредственно перед началом кембрия. . Можно также предположить, что вымирание само стало следствием предшествовавшего распада клатратов метана. Широко известно, что эмиссия метана и последующее насыщение атмосферы экосистеме не наблюдалось.
Ряд исследователей предполагал, что каждое кратковременное снижение доли 13 C/ 12 C в раннем кембрии представляет высвобождение метана, которое, благодаря вызванному им небольшому парниковому эффекту и повышению температуры, приводило к росту морфологического разнообразия. Но и эта гипотеза не объясняет резкого увеличения таксономического разнообразия в начале кембрия.
Объяснения на основе развития организмов
В основе ряда теорий лежит та идея, что относительно малые изменения в способе, которым животные развиваются из эмбриона во взрослый организм, могут привести к резким изменениям формы тела.
Отсюда видно, что наличие сходного набора Hox-генов вовсе не означает анатомического сходства организмов (раз одни и те же Hox-гены могут управлять формированием столь разных структур, как глаза человека и насекомого). Поэтому возникновение подобной системы могло повлечь резкий рост разнообразия - как морфологического, так и таксономического.
Поскольку одни и те же Hox-гены управляют дифференциацией всех известных билатеральных организмов, эволюционные линии последних должны были разойтись до того, как у них начали образовываться какие-либо специализированные органы. Таким образом, «последний общий предок» всех билатеральных организмов должен был быть небольшим, анатомически простым и, вероятнее всего, подверженным полному разложению без сохранения в окаменелостях. Это обстоятельство делает его обнаружение крайне маловероятным. Однако целый ряд вендобионтов (например, кимберелла Большая часть биологической сложности, вероятно, возникает благодаря действию относительно простых правил на большом числе клеток, функционирующих как клеточные автоматы (примером такого эффекта служит конвеевская игра «Жизнь» , где сложные формы и сложное поведение демонстрируются клетками, действующими по исключительно простым правилам). Возможное появление полового размножения или его существенное развитие в период кембрийского взрыва для очень примитивных и похожих существ может означать то, что существовала возможность их межвидового и более далёкого скрещивания. Это резко увеличивало изменчивость. Лишь с развитием генома появляются по настоящему изолированные виды, не скрещивающиеся с другими. Пример современных существ такого рода - кораллы .
Колея развития
Некоторые ученые предполагают, что по мере усложнения организмов, на эволюционные изменения общего строения тела накладываются вторичные изменения в сторону лучшей специализации его сложившихся частей. Это снижает вероятность прохождения естественного отбора новыми классами организмов - из-за конкуренции с «усовершенствованными» предками. В итоге, по мере складывания общего (на уровне таксономического класса) строения, формируется «колея развития», а пространственная структура тела «замораживается». Соответственно, формирование новых классов происходит «легче» на ранних стадиях эволюции основных клад , а их дальнейшая эволюция идет на более низких таксономических уровнях. Впоследствии автор этой идеи указывал, что такое «замораживание» не является основным объяснением кембрийского взрыва.
Окаменелости, которые могли бы подтвердить эту идею, неоднозначны. Отмечено, что вариации организмов одного класса зачастую наиболее велики на самых первых стадиях развития клады. Например, некоторые кембрийские трилобиты сильно варьировали по количеству грудных сегментов , причём впоследствии подобное разнообразие существенно снизилось. Однако обнаружено, что образцы силурийских трилобитов обладают столь же высокой вариативностью строения, что и раннекембрийские. Исследователи предположили, что общее снижение разнообразия связано с экологическими или функциональными ограничениями. Например, можно ожидать меньшей вариативности числа сегментов после того, как у трилобитов (напоминавших современную мокрицу) сформировалось выпуклое строение тела, являющееся эффективным способом его защиты.
Экологические объяснения
Такие объяснения сосредоточены на взаимодействии между различными видами организмов. Некоторые из подобных гипотез имеют дело с изменениями пищевых цепей; другие рассматривают гонку вооружений между хищниками и жертвами, которая могла вызвать эволюцию жестких частей тела в раннем кембрии; ещё какое-то число гипотез сосредоточено на более общих механизмах коэволюции (наиболее известен более поздний пример коэволюции цветковых растений с насекомыми-опылителями).
«Гонка вооружений» между хищниками и жертвами
Хищничество по определению предполагает гибель жертвы, в силу чего оно становится сильнейшим фактором и ускорителем естественного отбора. Давление на жертвы в направлении лучшей адаптации должно быть более сильным, чем на хищников - поскольку, в отличие от жертвы, они имеют шанс сделать новую попытку (эта асимметрия известна как принцип «жизнь против обеда» - хищник рискует потерей только обеда, в то время как жертва рискует жизнью). началось около 1,25 млрд лет назад - задолго до «взрыва».
Рост размеров и разнообразия планктона
Геохимические наблюдения четко показывают, что общая масса планктона стала сравнима с нынешней уже в раннем протерозое. Однако до кембрия планктон не вносил существенного вклада в питание глубоководных организмов, поскольку их тела были слишком малы для быстрого погружения на морское дно. Микроскопический планктон поедался другим планктоном или разрушался химическими процессами в верхних слоях моря задолго до проникновения в глубоководные слои, где мог бы стать пищей для нектона и бентоса (плавающие организмы и обитатели морского дна соответственно).
В составе же ранних кембрийских ископаемых был обнаружен мезозоопланктон (планктон средних размеров, различимый невооруженным глазом), который мог отфильтровывать микроскопический планктон (главным образом, фитопланктон - планктонную «растительность»). Новый мезозоопланктон мог служить источником останков, а также выделять экскременты в форме капсул, достаточно крупных для быстрого погружения - они могли быть пищей для нектона и бентоса, вызывая рост их размеров и разнообразия. Если же частицы органики достигали морского дна, в результате последующего захоронения они должны были повышать концентрацию кислорода в воде при одновременном снижении концентрации свободного углерода. Другими словами, появление мезозоопланктона обогатило глубокие участки океана как пищей, так и кислородом, и, тем самым, сделало возможным появление и эволюцию более крупных и разнообразных обитателей морских глубин.
Наконец, возникновение среди мезозоопланктона фитофагов могло сформировать дополнительную экологическую нишу для более крупных мезозоопланктонных хищников, чьи тела, погружаясь в море, вели к дальнейшему его обогащению пищей и кислородом. Возможно, первыми хищниками среди мезозоопланктона были личинки донных животных, чья дальнейшая эволюция стала результатом общего роста хищничества в морях эдиакарского периода.
Множество пустых ниш
Джеймс Валентайн (James W. Valentine) в нескольких работах сделал следующие предположения: резкие изменения в строении тела являются «затруднительными»; изменения имеют гораздо больше шансов на существование, если они встречают слабую (или вовсе не встречают) конкуренцию за ту экологическую нишу , на которую они нацелены. Последнее необходимо, чтобы новый тип организмов имел достаточно времени для адаптации к своей новой роли.
Это обстоятельство должно приводить к тому, что реализация основных эволюционных изменений гораздо более вероятна на начальных стадиях формирования экосистемы - из-за того, что последующая диверсификация заполняет почти все экологические ниши. В дальнейшем, несмотря на то, что новые типы организмов продолжают возникать, нехватка пустых ниш препятствует их распространению в экосистеме. . Москва, Наука, 1986.
Бизнес идеи